Глава 2. Физиология мужских половых органов (Михайличенко В.В.)
В функциональном отношении тесно связанные между собой половые органы мужчины подразделяются на: 1) половые железы - яички (семенники); 2) дополнительные половые образования (придаточные половые железы); 3) половые пути (семявыводящие пути); 4) органы совокупления.
Физиология яичек. Яички одновременно выполняют двойную функцию: герминативную и внутрисекреторную. Герминативная функция за счет сперматогенеза обеспечивает образование мужских половых клеток (спермиев), способствуя тем самым продолжению рода.
Внутрисекреторная функция заключается в выделении мужских половых гормонов (андрогенов), среди которых основным является тестостерон. Помимо андрогенов, в яичке образуются эстрогены, главным образом эстрадиол.
Тестостерон является наиболее активным андрогенным гормоном. Местом синтеза андрогенов у мужчин являются гландулоциты яичка (клетки Лейдига), расположенные в интерстициальной ткани яичек поодиночке или группами. Гландулоциты имеют значительные размеры, правильную форму и содержат в цитоплазме липоидные и пигментные включения. Тестостерон способствует возникновению вторичных половых признаков и либидо, созреванию мужских половых клеток - спермиев, - обладает выраженной анаболической активностью, стимулирует эритропоэз, существенно влияет на синтез белка, индуцирует ферменты. В больших дозах андрогены тормозят пролиферацию хрящевой ткани и стимулируют ее оссификацию; дефицит гормонов приводит к торможению процессов окостенения хряща. Под влиянием тестостерона, вырабатываемого яичками плода, происходят маскулинизация наружных и внутренних половых органов и развитие их по мужскому типу.
Средняя суточная продукция тестостерона в организме мужчин 25-40 лет колеблется, по данным О. Н. Савченко (1979), в пределах 4-7 мг.
Максимальная продукция андрогенов половыми железами наблюдается у мужчин в возрасте 25-30 лет, после этого начинается медленное снижение их гормональной активности. При старении уровень тестостерона в крови снижается, уровень эстрогенов - повышается.
На основании собственных исследований и изучения обширной литературы У. Мейнуоринг (1979) пришел к следующим выводам. Основной андроген (тестостерон) циркулирует с кровью в виде устойчивого комплекса с белками плазмы и подвергается интенсивному метаболизму только в клетках-мишенях для андрогенов. Основным его метаболитом является 5α-дегидротестостерон.
5α-дегидротестостерон представляет собой активный метаболит тестостерона, который с белками плазмы образует андрогенрецепторный комплекс, способный связываться с ядерными акцепторами и стимулировать многие биохимические процессы. Разрушение и вытеснение из ядра андрогенрецепторного комплекса приводит к замедлению основных биохимических процессов, обусловливающих андрогенный ответ.
Метаболизм тестостерона происходит под действием специального фермента 5α-редуктазы. В мужских добавочных половых железах содержится значительное количество 5α-редуктазы, при участии которой в них возможно образование 5α-дегидротестостерона. Было также установлено, что 5α-дегидротестостерон прочно связывается с ядрами клеток добавочных половых желез. В придаточных половых железах, мышцах и других тканях расположены клетки-мишени, которые являются акцепторами для тестостерона и его метаболитов и способны давать специфические андрогенные ответы.
Андрогены яичек плода вызывают регресс мюллеровых протоков и развитие из вольфовых протоков придатков яичек, семявыносящих протоков, семенных пузырьков, предстательной железы с маскулинизацией наружных половых органов.
Добавочные половые железы постоянно испытывают на себе влияние андрогенов, которые способствуют их правильному формированию и нормальному функционированию.
Тестостерон стимулирует образование фруктозы в семенных пузырьках, лимонной кислоты и фосфатазы в предстательной железе, корнитина в придатках яичка и пр.
Уменьшение содержания в семенной жидкости фруктозы, лимонной кислоты, кислой фосфатазы, корнитина может указывать на снижение внутрисекреторной функции яичек.
Обнаружено, что приблизительно через 7-10 дней после двусторонней орхиэктомии мужские добавочные половые железы у грызунов атрофируются до минимума. Последующее введение тестостерона приводит к значительному увеличению их массы и усилению внутриклеточной секреции.
Таким образом, биологические ответы на андрогены направлены на поддержание структуры и функции андрогенных клеток-мишеней, типичным примером которых являются клетки мужских добавочных половых желез.
Изучение механизма действия гормонов осложняется взаимопревращением андрогенов в эстрогены и андростендиола (основного андрогеноподобного стероида, секретируемого надпочечниками) в тестостерон.
В настоящее время не вызывает сомнений, что некоторые биохимические явления специфически регулируются самим тестостероном, другими активными метаболитами и даже эстрогенами.
80% эстрогенов у мужчин образуется в яичках и лишь 20% - в надпочечниках. Биологическое значение эстрогенов в мужском организме заключается в стимулирующем влиянии на интерстициальные клетки половых желез, гладкую мускулатуру, соединительную ткань и специфический эпителий.
Большое значение в организме человека придается антиандрогенам. У. Мейнуоринг (1979) отмечает антиандрогенное действие эстрогенов, основанное на подавлении секреции гонадотропинов, ингибировании системы 5α-редуктазы и стимуляции синтеза половых стероидных гормонов. В какой-то степени эстрадиол может конкурировать с 5α-дегидротестостероном за связывающие места, но только в том случае, если присутствует в избытке.
Андрогенные стероиды вырабатываются как яичками, так и надпочечниками. "Корой надпочечников вырабатываются производные андростана, обладающие андрогенной активностью: 17-кетостероиды (дегидроэпиандростерон, этиохоланолон, андростендион, андростерон) - и мужской половой гормон тестостерон, а также производные эстрана - эстрогены (эстрадиол и эстрон). Важным промежуточным продуктом синтеза гормонов коры надпочечников является прогестерон. Значительная часть андрогенов, претерпевая изменения, выделяется почками в виде нейтральных 17-кетостероидов (17-КС).
Из общего количества 17-КС, выделяемых с мочой, 1/3 образуется за счет метаболизма соединений, продуцируемых клетками Лейдига яичка, а 2/3 - клетками коры надпочечников. Понятно, что колебания уровня экскреции 17-КС зависят от состояния ЦНС и системы гипоталамус - гипофиз - надпочечники. На самом деле определение 17-КС в моче дает лишь общие сведения о метаболизме стероидных соединений, вырабатываемых и яичками, и корой надпочечников. Поэтому определение выделения 17-КС с мочой не может служить методом оценки эндокринной функции гландулоцитов яичка.
Таким образом, становится понятно, что только прямое определение тестостерона и эстрадиола в крови и моче, являющихся в основном продуктами яичек (в мужском организме), может служить показателем гормональной функции яичек.
Одной из важнейших функций андрогенов и в особенности тестостерона является поддержание процесса сперматогенеза. Состояние сперматогенеза зависит от концентрации андрогенов в тестикулярной ткани, и потому снижение образования тестостерона может быть одной из основных причин неплодовитости мужчин.
Для полного течения процесса сперматогенеза важна также роль андроген-связывающего белка, который образуется в яичках и способствует переносу андрогенов в цитоплазму клеток сперматогенного эпителия. Цитоплазматический рецептор, соединяясь с андрогенами, облегчает их проникновение непосредственно в ядра.
Сперматогенез. Процесс сперматогенеза осуществляется в извитых семенных канальцах паренхимы яичка, которые составляют основную объемную массу его. Внутренняя поверхность мембран извитых канальцев выстлана двумя видами клеток, сустентоцитами и первичными половыми клетками - сперматогониями. Именно здесь недифференцированные семенные клетки сперматогонии размножаются и превращаются в зрелые спермии.
В течение эмбрионального развития и в детском возрасте первичные сперматогонии делятся митотически, давая начало дополнительным сперматогониям. С 10-летнего возраста в семенных канальцах мальчиков начинаются усиленное митотическое деление сперматогоний и формирование сустентоцитов. Начальные этапы сперматогенеза появляются в 12-летнем возрасте - происходит образование сперматид из сперматоцитов второго порядка. Полное формирование сперматогенеза происходит к 16 годам.
Изнутри мембрану семенного канальца выстилают сустентоциты, которые обеспечивают сперматогенные клетки продуктами своей секреторной активности, выполняют фагоцитарную функцию в отношении остатков после сперматогенеза, синтезируют эстрогеноподобное вещество (ингибин), выделяют андрогенсвязывающий протеин, способствующий переносу тестостерона и дигидротестостерона в половые клетки, где они закрепляются в ядре, обусловливая различные метаболические процессы, необходимые для созревания спермиев. Как бы втиснутые между сустентоцитами, ближе к основанию мембраны располагаются сперматогонии. В просвет канальца направлены многочисленные цитоплазматические отростки сустентоцитов, между отростками располагаются клетки сперматогенного эпителия. По мере созревания клетки сперматогенного эпителия продвигаются к просвету канальца. В результате митотического деления приумножается число сперматогоний. Последние, увеличиваясь в размерах, превращаются в сперматоциты первого порядка, каждый из которых содержит диплоидный набор хромосом 46XY. Сперматоциты первого порядка после усиленного роста и созревания вступают в стадию мейоза (редукционного деления). При этом из сперматоцитов первого порядка образуются 2 сперматоцита второго порядка с гаплоидным набором хромосом (22 аутосомы и 1 половая - X или Y). Из каждого сперматоцита второго порядка образуются путем быстрого митотического деления 2 сперматиды. В конечном итоге из одного сперматоцита первого порядка образуются четыре сперматиды, содержащие уменьшенный наполовину гаплоидный набор хромосом. Сперматиды захватываются цитоплазматическими выростами сустентоцитов, в цитоплазме которых происходит развитие и формирование спермиев. Сперматида удлиняется, ядро ее смещается эксцентрично. Из части цитоплазмы формируется шейка и вырастает жгутик спермия. После распада протоплазменных выростов сустентоцитов спермии освобождаются и выходят в просвет канальцев, накапливаются в придатке, где происходит их дозревание.
Развитие и дифференциация спермиев проходят 3 этапа: 1) пролиферация сперматогоний - сперматоцитогенез, 2) деление и созревание сперматоцитов - сперматогенез и 3) конечная фаза, дифференциации сперматид в спермии - спермиогенез. Профаза первого (мейотического) деления сперматоцита 1-го порядка занимает значительную долю (около 3/8) времени сперматогенеза. Второе (митотическое) деление сперматоцитов 2-го порядка, приводящее к образованию сперматид, происходит довольно быстро. Морфологические изменения в сперматиде, включающие в себя перестройку ядра и цитоплазматических элементов и завершающиеся образованием спермиев, суммарно описываются как спермиогенез и также длятся около 3/8 времени сперматогенеза (рис. 4). Время, необходимое для превращения первичной клетки в спермий, занимает у человека около 74-75 дней. Жидкость, заполняющая просвет семенных канальцев, представляет собой продукт секреции клеток семенных канальцев и содержит гормон (ингибин), который угнетает продукцию фолликулостимулирующего гормона (ФСГ) гипофизом. При поражении семенных канальцев и угнетении сперматогенеза уменьшается продукция ингибина, это приводит к повышенному выделению гонадотропинов гипофизом.
В зародышевом эпителии яичек образуется фермент гиалуронидаза, которая локализуется в головке спермиев. Небольшое количество гиалуронидазы попадает в плазму эякулята из спермиев. Гиалуронидаза растворяет слизь шейки матки и обладает свойством разъединять клетки лучистого венца (corona radiata) яйцеклетки без их деструкции и таким образом создавать возможность для внедрения в нее спермия. Значительная концентрация гиалуронидазы создается достаточным количеством спермиев. При аспермии гиалуронидаза в эякуляте отсутствует.
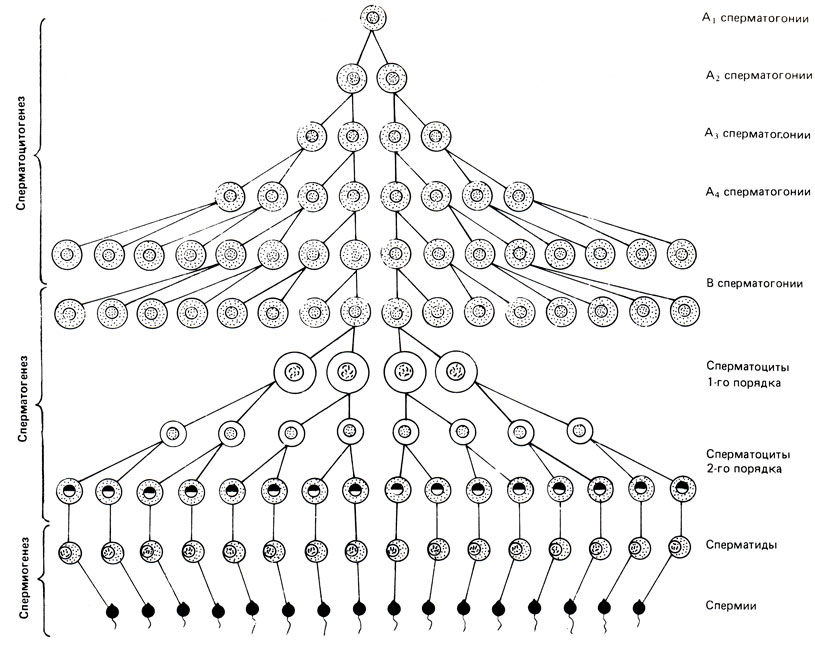
Рис. 4. Схема сперматогенеза
Другим продуктом секреции яичек являются простагландины, открытые шведским ученым Euler в 1936 г. Предполагалось, что они образуются в предстательной железе. Затем было установлено, что основным местом их образования являются яички. Было доказано влияние простагландинов на сократительную способность гладкой мускулатуры и стимулирующее влияние на выработку ФСГ и ЛГ. Из выделенных в настоящее время нескольких десятков простагландинов практическое значение имеют два вида: Е2 - очень неустойчивый и Е2α - стойкий. В больших объемах эякулята содержится большее количество простагландинов. Их способность расслаблять и сокращать гладкую мускулатуру женских половых путей повышает скорость прохождения яйцеклетки через маточные трубы навстречу спермиям в процессе зачатия. Высокое содержание простагландинов стимулирует сократительную способность гладкой мускулатуры матки, прерывая беременность.
Базальная мембрана канальцев (особенно мышечноподобные клетки внутреннего слоя и сустентоциты) создают гематотестикулярный барьер, защищающий генеративный эпителий, ответственный за наследственность и продление рода, от инфекционных и токсических поражений.
Исследование эякулята позволяет судить о степени и характере нарушений внутрисекреторной и экскреторной функций яичек, так как тестостерон и гонадотропины влияют на морфологические и физико-химические свойства эякулята.
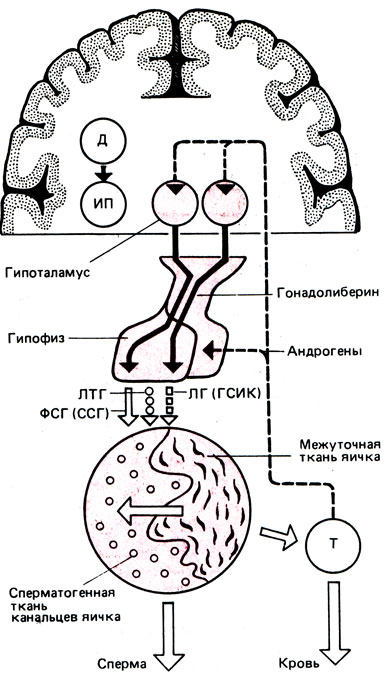
Рис. 5. Регуляция функции яичек. Д - дофамин; ИП - ингибитор пролактина; Т - тестостерон
Регуляция функции яичек. Деятельность яичек находится непосредственно под влиянием ЦНС, гипоталамуса и гипофиза. Кора головного мозга выполняет наиболее ответственную функцию - приспособление деятельности эндокринной системы к постоянно изменяющимся факторам внешней и внутренней среды. Действие коры головного мозга на половые железы реализуется через гипоталамус - гипофиз или через изменение функционального состояния вегетативной нервной системы, приводящее к расстройствам кровообращения. Надо полагать, что, наряду с нарушением васкуляризации, нарушается обмен веществ в иннервируемом органе (яичках), что и приводит к нарушению сперматогенеза.
Роль нервной системы и гипоталамических центров при регулировании функций мужских половых желез заключается в их влиянии, осуществляемом не только нейрогенным путем, но и через секрецию гипофиза, гормоны которого стимулируют функцию яичек. Гормоны, выделенные нервными клетками и определенными ядрами гипоталамуса, доставляются к гипофизу и стимулируют выделение гонадотропных гормонов.
Гипоталамус и гипофиз следует рассматривать как комплекс двух тесно связанных желез внутренней секреции (рис. 5). Рилизинг-гормон, вырабатываемый гипоталамусом, оказывает прямое влияние на стимуляцию или торможение секреции гормонов гипофиза. Выработка гонадотропин-рилизинг-гормона происходит преимущественно в области аркуатных ядер и стимулируется дофамином. Серотонин, выделяемый эпифизом, тормозит продукцию рилизинг-гормона. У мужчин функционирует постоянный тонический центр секреции рилизинг-гормона, у женщин - циклический. Такая половая дифференцировка гипоталамуса происходит во внутриутробном периоде под влиянием вырабатываемого эмбриональными яичками тестостерона.
В настоящее время установлено, что синтез и выделение гонадотропных гормонов регулируются единым гонадотропин-рилизинг-гормоном. М. Ammos и A. Sehally (1971) осуществили его синтез. Передняя доля гипофиза секретирует 3 гонадотропных гормона, которые оказывают влияние на функцию семенников.
ФСГ, именуемый в мужском организме сперматогенезстимулирующим гормоном (ССГ), активно влияет на сперматогенез, стимулирует эпителий канальцев яичка. ЛГ у мужчин инициирует развитие, созревание интерстициальных клеток и влияет на биосинтез андрогенов, поэтому носит название гормона, стимулирующего интерстициальные клетки (ГСИК).
Роль третьего гормона - пролактина, или лютеотропного гормона (ЛТГ), - в мужском организме долгое время оставалась неизвестной. Исследования последних лет показали, что пролактин является гормоном широкого спектра действия, в том числе и регулятором половой функции у мужчин. Пролактин потенцирует действие ЛГ и ФСГ, направленное на восстановление и поддержание сперматогенеза, увеличивает массу семенников и семенных канальцев, под влиянием пролактина усиливаются обменные процессы в яичке. Совместное назначение ЛГ и пролактина значительно больше повышает содержание тестостерона в плазме крови, чем при назначении только ЛГ. Пролактин подавляет образование дегидротестостерона.
Подавление образования дегидротестостерона из тестостерона в предстательной железе под влиянием пролактина осуществляется за счет торможения активности 5α-редуктазы. Изменяя таким образом обмен андрогенов, пролактин стимулирует секрецию предстательной железы по сравнению с ее ростом. У человека отчетливо прослеживается зависимость между содержанием пролактина в эякуляте и числом подвижных спермиев. В зависимости от степени снижения концентрации пролактина отмечаются низкая подвижность спермиев, олиго- или азооспермия. Процесс сперматогенеза у человека и животных прекращается после выключения гипофиза. В таких случаях сперматогенез блокируется уже на этапе сперматоцитов 1-го порядка еще до редукционного деления. Считают, что ФСГ стимулирует рост семенных канальцев, функцию клеток Сертоли, инициирует митотическую фазу сперматогенеза (от сперматогоний до сперматоцитов). Под влиянием ЛГ функционируют клетки Лейдига, вырабатывая тестостерон, который обеспечивает заключительную фазу сперматогенеза (спермиогенез) - превращение сперматоцитов в сперматиды и созревание их в спермии (рис. 6).
С другой стороны, андрогены действуют на диэнцефальную область, оказывают стимулирующее действие также на высшие корковые центры. При этом усиливаются положительные условные рефлексы и повышается тонус коры головного мозга.
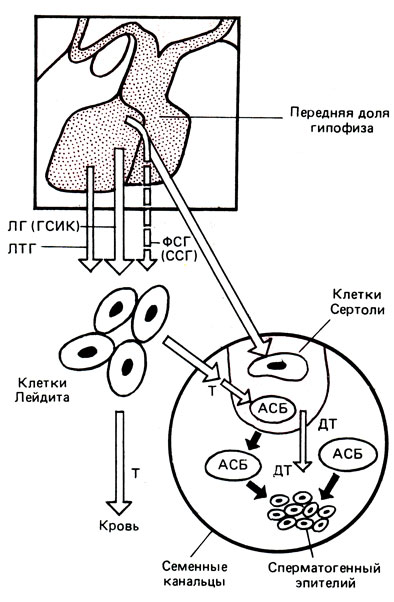
Рис. 6. Регуляция сперматогенеза. Т - тестостерон; ДТ - дигидротестостерон; АСБ - андрогенсвязывающий белок
Андрогены и эстрогены при длительном введении в больших дозах приводят к торможению нейросекреции гипоталамуса, к исчезновению гонадотропинов и к расстройствам сперматогенеза. Разрушение рецепторной (для половых стероидов) зоны гипоталамуса приводит к состоянию, имитирующему посткастрационное, что объясняется выключением афферентного звена в механизме обратной связи. Это свидетельствует о том, что местом приложения половых стероидов является передний гипоталамус, и объясняет также механизм развития бесплодия при некоторых диэнцефальных поражениях. Секреция ФСГ регулируется частично некоторыми неандрогенными факторами, имеющими неспецифическую связь со сперматогенезом, и частично тестостероном и его метаболитами. Поэтому при тяжелом, нарушении сперматогенеза вследствие крипторхизма наблюдается повышение уровня ФСГ в сыворотке крови. Установлена обоюдная количественная корреляция между уровнями ФСГ и тестостерона, что указывает на отрицательную регуляцию по типу обратной связи, существующей между ФСГ и тестостероном. Нарушение обратной чувствительности гипоталамоипофизарной системы к уровню периферического тестостерона (имеющее место при синдроме Клайнфелтера), чрезмерное повышение секреции гонадотропина приводят к вторичным изменениям гландулоцитов яичка и снижению уровня тестостерона. Таким образом, существование автоматической регуляции гипоталамической деятельности гипофиза и яичек является важным биологическим процессом, управляющим сперматогенной и инкреторной функцией половых желез.
Придаток яичка - представляет собой андрогензависимый, секреторно-активный орган, который служит для проведения накопления и дозревания спермиев. В придатке яичка под действием андрогенов создается благоприятная среда для завершения их развития и жизнедеятельности. По мере продвижения от головки до хвоста, продолжающегося в норме 14 дней, происходит окончательное морфологическое, биохимическое и физиологическое созревание спермиев, обретающих способность двигаться и оплодотворять яйцеклетку. В придатке спермин освобождаются от цитоплазматической капли (остатки цитоплазмы сустентоцитов), обволакиваются защитной белковой оболочкой, приобретают отрицательный заряд и насыщаются секретом, содержащим гликоген, жиры, холестерин, фосфаты, корнитин и др., происходит ряд ультраструктурных и цитохимических преобразований акросомы. По мере продвижения и созревания спермии накапливаются в хвосте, который является для них хранилищем. Концентрация спермиев здесь может быть в 10 раз больше, чем в нормальном эякуляте. Слабое напряжение кислорода и отсутствие фруктозы препятствуют активному метаболизму в спермиях и способствуют длительному сохранению их жизнедеятельности. При половом воздержании в хвосте придатка могут обнаруживаться и старые, переродившиеся формы спермиев. Эпителий придатка способен дезинтегрировать и абсорбировать нежизнеспособные формы спермиев. В этом процессе значительную роль играют спермиофаги. Способность к поглощению и усвоению спермией создает условия для поддержания сперматогенеза у больных с обструктивной аспермией, при частичном сохранении функционирующей части придатка. При полном поражении придатка яичка сперматогенез нарушается, так как наступают переполнение и гибель канальцев яичка. Продвижение спермиев из яичек в придаток и в самом придатке осуществляется за счет движения ресничек мерцательного эпителия выносящих канальцев и давлением непрерывно поступающего секрета яичка.
Семявыносящий проток является органом, служащим для проведения спермиев от хвоста придатка до ампулы семявыносящего протока, где происходит их накопление. При половом возбуждении спермии могут накапливаться также и на длинном отрезке между ампулой и хвостовой частью придатка яичка. При эякуляции опорожняются прежде всего ампула и периферический отрезок семявыносящего протока. Содержимое семявыносящего протока проталкивается во время эякуляции по направлению к уретре за счет укорочения всего придатка в результате сокращения его мощной мускулатуры. При последующих извержениях количество спермиев значительно уменьшается и поступают они из хвоста придатка яичка, который полностью никогда не опорожняется.
Семенные пузырьки являются железистыми андрогензависимыми секреторными органами. Секрет семенных пузырьков состоит из вязкой, беловато-серой желатиноподобной субстанции, которая после эякуляции разжижается в течение нескольких минут и составляет около 50-60% семенной жидкости. Наиболее важной функцией семенных пузырьков является секреция фруктозы, уровень которой является показателем андрогенной насыщенности организма. Фруктоза служит источником энергии, метаболизма и поддержания подвижности спермиев. Нормальное содержание фруктозы в сперме здорового мужчины 13-15 ммоль/л. При хранении эякулята количество фруктозы уменьшается из-за потребления ее спермиями. Расходование фруктозы спермиями (фруктолиз) в нормальном эякуляте не ниже 3-5 ммоль/л за 2 ч. Семенные пузырьки секретируют и другие составные части спермы: азотистые вещества, белки, инозит, аскорбиновую кислоту, простагландины и др. Секрет семенных пузырьков с рН 7,3, примешиваясь к секрету яичек, играет роль защитного коллоида, придавая спермиям большую сопротивляемость. При нереализованном половом возбуждении спермии попадают в семенные пузырьки, где они могут поглощаться клетками-спермиофагами. Семенные пузырьки также способны к резорбции жидких компонентов.
Предстательная железа - андрогензависимый орган, поставляющий около 25-35% плазмы спермы. При уменьшении уровня андрогенов в крови ее секреторная активность значительно снижается. Слабощелочной секрет предстательной железы в норме содержит значительное количество преломляющих свет зернышек (липоидных телец), которые придают ему опалесцирующий беловатый оттенок. Значительное содержание в секрете предстательной железы спермина придает эякуляту характерный запах. При медленном охлаждении в эякуляте появляются кристаллы фосфата спермина. Фибринолизин и фиброгеназа, являясь мощными протеолитическими ферментами, принимают участие в разжижении эякулята.
В предстательной железе также образуется лимонная кислота, концентрация которой служит показателем функционального состояния ее и своеобразным "андрологическим эквивалентом" эндокринной функции яичек.
В норме концентрация лимонной кислоты в семенной жидкости колеблется от 2,5 до 3,5 ммоль/л. Секрет предстательной железы содержит кислую и щелочную фосфатазы. Отношение содержания кислой фосфатазы к щелочной (фосфатазный индекс) является довольно стабильной величиной [Юнда И. Ф., 1982]. Под действием фосфатазы происходит расщепление холин-фосфорной кислоты семенной плазмы на холин и фосфорную кислоту. Спермин, соединяясь с фосфорной кислотой, образует кристаллы фосфорного спермина. Холин оказывает сенсибилизирующее действие на клетки. Спермин и спермидин, являясь основаниями, поддерживают на постоянном уровне концентрацию водородных ионов. В предстательной железе вырабатываются простагландины, влияющие на сократительную деятельность гладкой мускулатуры. Высказывается мнение об эндокринной функции предстательной железы. Однако убедительные данные в пользу этого до сих пор отсутствуют. В клетках-мишенях предстательной железы происходит метаболизм тестостерона. Под действием 5α-редуктазы тестостерон превращается в еще более активный метаболит 5α-дегидротестостерон, способный с белками плазмы образовывать андрогенрецепторный комплекс, который может проникать в ядерные структуры и стимулировать многие биохимические процессы.
Приведенные данные показывают, что предстательная железа увеличивает объем эякулята, участвует в разжижении, оказывает буферное и ферментативное действие на эякулят в целом и активизирует движение спермиев. Функционально предстательная железа тесно связана с семявыводящими путями. Патологические изменения в ней могут приводить к нарушениям репродуктивной и копулятивной функций. Размеры предстательной железы значительно меняются с возрастом. Железистая ткань ее развивается в период полового созревания и дегенерирует у стариков.
Луковично-уретральные железы являются гомологом бартолиновых желез. Секрет этих желез, выделяющийся в мочеиспускательный канал при половом возбуждении за счет сокращения мышц промежности, представляет собой бесцветную прозрачную, лишенную запаха слизь со щелочной реакцией. При прохождении по уретре секрет нейтрализует кислую реакцию остающейся в ней мочи и, выделяясь из наружного отверстия мочеиспускательного канала, облегчает введение полового члена во влагалище. С возрастом отмечается гипотрофия луковично-уретральных желез.
Уретральные железы. На всем протяжении слизистой оболочки мочеиспускательного канала, особенно на передней и боковых стенках его, располагаются гроздьевидные, трубчато-альвеолярные парауретральные железы, выделяющие слизистый секрет, количество которого увеличивается при половом возбуждении. Он служит для увлажнения уретры и вместе с секретом луковично-уретральных желез поддерживает щелочную реакцию, благоприятную для спермиев.
Семенной холмик - представляет собой возвышение на задней стенке предстательной части уретры, в середине которого располагается мужская маточка - рудимент мюллеровых протоков. Длина маточки - около 8-10 мм. В центре маточки, которая внедряется в толщу вещества предстательной железы, открывается щель, переходящая в неглубокую (до 4-6 мм) полость. На дне этой полости или ниже ее на семенном холмике открываются щелевидные устья семявыбрасывающих протоков. Семенной холмик состоит из пещеристой ткани, богатой эластическими волокнами и продольными пучками гладких мышц. По бокам семенного бугорка открываются (по 10-12 с каждой стороны)' устья выводных протоков долек предстательной железы.
Физиологическое значение семенного бугорка до конца не изучено. Будучи эмбриологически и анатомически связанным с органами половой системы, семенной бугорок активно участвует в акте эякуляции. Вокруг него концентрируются выводные протоки большинства половых желез и нервные окончания, связанные с центром эякуляции.
Мочеиспускательный канал у мужчин изменяется с возрастом. До периода полового созревания канал короче, уже и имеет резкий изгиб в заднем отделе. После полового созревания в связи с увеличением полового члена, развитием предстательной железы уретра формируется окончательно. В пожилом возрасте при гипертрофии парауретральных желез изменяется предстательный отдел мочеиспускательного канала и уменьшается его просвет.
Мочеиспускательный канал выполняет 3 функции: удерживает мочу в мочевом пузыре; проводит мочу при мочеиспускании; проводит семенную жидкость в момент эякуляции. Удержание в мочевом пузыре мочи осуществляется за счет внутреннего (непроизвольного) и наружного (произвольного) сфинктеров. При переполнении мочевого пузыря основную роль играет мощный наружный произвольный сфинктер, удерживать мочу помогает и сокращение мышечной массы предстательной железы. Мочеиспускание является сложным рефлекторно-произвольным актом. Когда внутрипузырное давление достигает определенного уровня (при объеме мочи в мочевом пузыре свыше 200 мл), появляется позыв к мочеиспусканию. Под влиянием волевого импульса происходит сокращение мышц мочевого пузыря и брюшной стенки с одновременным расслаблением сфинктеров, и мочевой пузырь опорожняется.
Проведение семенной жидкости по мочеиспускательному каналу осуществляется в момент эякуляции. Эякуляция - рефлекторный акт, в котором активное участие принимают сам мочеиспускательный канал и все образования, связанные с ним. При этом происходит сокращение внутреннего сфинктера, который вместе с набухшим во время эрекции семенным холмиком препятствует забрасыванию эякулята в мочевой пузырь. Одновременно с этим расслабляется наружный сфинктер и происходит последовательное опорожнение от содержимого придатков яичек, семявыносящих протоков, включая ампулярную часть, после чего возникает сокращение гладкой мускулатуры семенных пузырьков и предстательной железы, присоединяются мощные сокращения поперечнополосатой мускулатуры, седалищно-пещеристых и пещеристо-луковичных мышц и мышц тазового дна и промежности, в результате чего эякулят выбрасывается наружу со значительной силой. Регуляция акта эякуляции осуществляется симпатическим и парасимпатическим отделами нервной системы и под воздействием импульсов, следующих из ThXII-LIV и SII-IV сегментов спинного мозга.
Половой член является органом, который способен при возбуждении увеличиваться и приобретать значительную плотность, что необходимо для введения его во влагалище, совершения фрикций и подведения эякулята к шейке матки. В состоянии эрекции головка полового члена остается эластичной, что препятствует травмированию женских половых органов. Эрекция - рефлекторный акт, в основе которого лежит наполнение кровью кавернозных тел, имеющих многокамерное сетчатое строение. Г. Вагнер (1985) выделяет 4 фазы эрекции.
Фаза покоя характеризуется постоянным объемом полового члена, внутрипещеристым давлением и объемом крови в половом члене. В этом состоянии внутрипещеристое давление равно около 5 мм рт. ст., объем оттекающей крови - от 2,5 до 8 мл/мин и равен объему притекающей.
Фаза набухания проявляется увеличением объема полового члена, сопровождающимся постепенным возрастанием внутрипещеристого давления до 80-90 мм рт. ст. Продолжительность ее зависит от интенсивности сексуальной стимуляции, восприимчивости к ней и возраста мужины. При этом увеличивается приток артериальной крови до 90 мл/мин, а отток остается прежним.
Фаза эрекции определяется постоянным объемом напряженного полового члена, увеличением внутрипещеристого давления по меньшей мере до 80 мм рт. ст., достигающего уровня артериального.
Объем артериального притока крови в период наступления эрекции составляет от 120 до 270 мл/мин.
Фаза детумесценции проявляется исчезновением ригидности полового члена и уменьшением объема с постепенным возвращением к исходному уровню. Это достигается за счет резкого увеличения оттока крови до 40 мл/мин, одновременно с этим постепенно уменьшается приток и снижается внутрипещеристое давление.
В период набухания полового члена сохраняется отток крови по системе дорсальной вены, но увеличивается приток артериальной крови. В пожилом возрасте период набухания удлиняется, что, очевидно, объясняется ухудшением притока артериальной крови. Во время эрекции отток крови по системе дорсальной вены прекращается, вновь появляясь лишь при высоком внутрипещеристом давлении, и восстанавливается полностью после эякуляции в фазе детумесценции. Тем не менее во время эрекции ток крови сохраняется, что обеспечивает ее достаточную длительность при нереализованном половом акте. Эрекция регулируется при помощи парасимпатических волокон, идущих в составе n. erigentes, импульсами их крестцовых и спинномозговых центров, находящихся под контролем высших нервных центров коры головного мозга.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://sohmet.ru/ 'Sohmet.ru: Библиотека по медицине'